Research
More than a century’s worth of behavioural investigations have demonstrated that animals and humans
process sensory information close to optimally, often employing subtle and powerful algorithms to
do so. Our understanding of these computations at the neural level is, by contrast, quite simplistic.
The goal of research in my group is to help bridge this gap, using both theoretical and data-driven
approaches to understand how information is represented in neural systems, and how this representation
underlies computation and learning. On the one hand, we collaborate closely with physiologists to
advance the technology of neural data collection and analysis. These studies have the potential to
introduce powerful new theoretically-motivated ways of looking at neural data. At the same time, we
examine neural information representation and perceptual behaviour from a more theoretical point
of view, addressing questions of how the brain might encode the richness of information needed to
explain perceptual capabilities, what purpose might be served by adaptation in neural activities, and
how experience-driven plasticity in representations is related to perceptual learning. Both the data
analytic and the theoretical aspects of our neuroscience research are closely connected to the field of
machine learning, which provides the tools needed for the first, and a structural framework for the
second.
Current and Recent Projects
Theoretical Neuroscience
Theoretical research in my group is built around a key idea in modern theoretical neuroscience: that the act of
perception is a search for the causal elements most likely to account for the activity of the sensory epithelia. This
”statistical inference” hypothesis, traceable to the work of von Helmholtz and before, has far-reaching consequences
for the way we study and think about brain function, and work in my group has explored many of its different
facets.
- The statistics of the sensory environment
If perception does indeed proceed by statistical inference, the brain must embody knowledge of the
statistical causal structure of the natural environment. We seek to discover this structure, using the
methods of probabilistic unsupervised learning to build statistical models of natural stimuli, and
drawing parallels between the elements of such models and neural processing and behaviour.
- Many statistical models of image data have relied on sparse models of static images or “slow”
models of video. We have taken a probabilistic approach to both sparsity [33] and slowness [45];
and have synthesised them in a structured model for natural image sequences [25], which matches
both the receptive field properties and anatomical grouping of cells in the visual cortex, and
suggests a new interpretation of complex and simple cells properties as representing the presence
and appearance respectively of visual features. We have also explore sparse image models with
competitive or occlusive source interactions, rather than the conventional linear superposition.
[26, 37, 43]. (With Pietro Berkes, Jörg Lücke, Richard Turner)
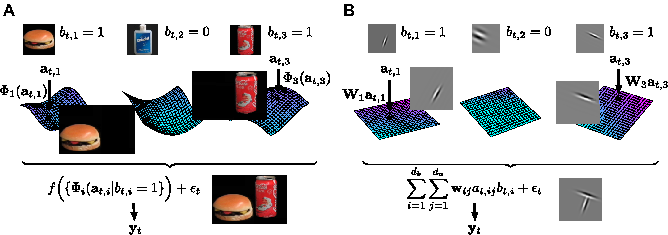
Identity-Attribute models. A natural image can be characterised by a set of
binary variables that indicate which objects appear, and a set of variables
specifying attributes such as apparent size and rotation, which determine
how they look. In the real world (left), this leads to a substantially nonlinear
model. In our simplification (right, [25]) these nonlinearities are reduced to
a single product. Despite this, the model captures essential elements of the
structure of video.
- Similar ideas should extend to sound, but here time must play an even more crucial role. We have studied
the statistics of sound in both signal-processing [23, 44] and more natural [39] settings. Much of our work
relies on the statistics of processes formed by a hierarchical cascade of modulators (With Richard
Turner).
- Perception
Much of our work focuses directly on perception, building probabilistic models of specific perceptual
processes—in some cases based on the statistical models derived above—and using these to understand human
and animal behaviour. In many cases, these models make behavioural or psychophysical predictions that we
are able to test experimentally.
- Other themes
We also have interests in the representation and manipulation of uncertainty by neural populations [58];
task-driven plasticity in primary sensory cortex [51]; sensory adaptation [55]; statistical outlier detection in the
nervous system; and motor planning (in collaboration with the Neural Prosthetic Systems Laboratory at
Stanford University).
Neural Data Modelling
Ultimately, any attempt to link theory to neural circuits will demand a sophisticated understanding of both
representation and dynamics within real neural systems. Thus, the group maintains substantial collaborations with
neurophysiologists. Our goals in these collaborations are both technological and scientific—developing the
algorithms needed to make sense of the flood of neurophysiological data available, while at the same time gaining
insight into representations and computations in the brain.
- Dynamics in neural populations
Neurons coöperate in large interconnected populations to represent and process information. Technical
limitations over the last many decades, however, have meant that almost all of what we know about
neuronal processing has been based on inferences made from the activity of cells measured one at a
time. Over the last five to ten years, the technology to monitor simultaneous activity in larger ensembles
has been more widely adopted, and this new stream of data provides new opportunities and poses new
challenges.
Our approach to such data (in collaboration with Krishna Shenoy’s laboratory at Stanford and other
groups in the DARPA REPAIR programme; [42]) rests on the idea the the collective activity of
multiple cells in the same area can be described by the evolution of a smaller set of latent variables
or “order parameters” that capture the dynamics of the network. We have worked on parametric
techniques to identify linear or non-linear dynamics in such systems [13, 14, 47, 48], and more recently
on Gaussian Process methods to identify underlying latent trajectories without constrained dynamics
[28, 29, 35, 36]. Our approach has begun to yield new insight into the process of motor preparation
[10] and dynamics within many different cortical areas [20]. (With Jakob Macke, Biljana Petreska, Alex
Lerchner, Lars Büsing, Byron Yu, John Cunningham, Afsheen Afshar, Krishna Shenoy, and others at
Stanford. )
- Neural encoding
A substantial current focus in auditory research is on understanding representation at the first cortical
stages of the auditory pathway. By building models of how cells in the primary auditory cortex
respond to stimuli, Jennifer Linden (UCL Ear Institute) and I hope to understand the function of
these cortical areas, and how that function is affected by acoustic context and auditory learning. In
early work we looked at the standard models for central auditory responses—called spectro-temporal
receptive fields (STRFs)—and developed methods both to improve the fidelity with which they might
be estimated, and to measure their predictive quality [59, 60]. The conclusion was that the best-fit
model could capture no more than 30% of the predictable variance in the response. This discovery
has led us to question the intepretation of STRF models [34]; and to build new nonlinear models
that improve predictive performance [30, 31]. These latter models have now led to exciting new
results, demonstrating that different cortical subfields have distinct nonlinear properties, and that
auditory learning is associated with changes in nonlinear cortical processing. (With Misha Ahrens,
Ross Williamson, Bjorn Christianson, and Jennifer Linden).
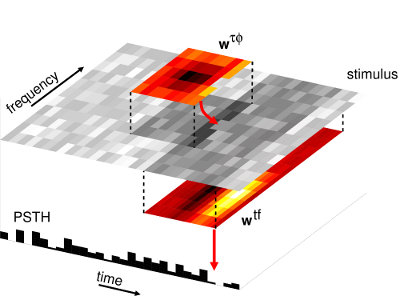
The context model. This multilinear model [30] captures short-term context effects in
the neural response to complex sounds.
As simultaneous recordings from multiple neurons become more commonplace, we see the emphasis in
experimental encoding studies moving from single cells to population-based methods. We have already begun
to gather such multi-site data from the auditory cortex, and have also initiated new collaborations with two
groups at UCL that are already collecting remarkably rich data sets in the visual cortex. Our
collaboration with Matteo Carandini has begun by looking at interactions between neurons and
local field potential recordings at sites regularly spaced across the visual cortex. At the same
time, with Tom Mrsic-Flogel, we are beginning to look at calcium imaging data from a number
of individual cells packed into a relatively small cortical region. In both cases, we hope to be
able to use techniques and algorithms developed as part of our population dynamics models in
motor areas to advance our understanding of sensory processing. (With Jakob Macke, David
Schulz)
- Neural decoding
At the other end of sensory-motor processing, the question of how neural activity encodes intended or current
movements—and how this activity may be decoded to provide effective motor facilities in paralysed
patients—is of considerable interest. Collaborating with the Shenoy group again, we have developed
probabilistic methods for decoding [40], with a particular emphasis on combining information about both
intended and current motor actions [24, 27, 38, 46, 56]. These algorithms are being implemented in real-time
systems, with a view to clinical development, as part of the DARPA Revolutionising Prosthetics 2009
programme.
Machine Learning and Signal Processing
At the heart of our work lie the methods of probabilistic (or Bayesian) modelling, particularly as they apply to the
fields of Machine Perception and Learning, and Signal Processing. Some of our work is directed more specifically at
understanding and developing these tools.
- Approximation
We are particularly interested in characterising the impact of the sorts of deterministic approximation
often used to accelerate inference in Machine Learning applications. Particularly in time series, common
approaches such as Laplace approximation, variational methods and EP introduce potential biases
[17, 47], some of which may be counter-intuitive. As such methods have become a central element of
the modelling toolbox, understanding these biases is of substantial importance.
- Sparsity
Models with sparse priors play a major role in modern signal processing. We have developed a variety
of probabilistic approaches to sparsity; exploiting hyperparameter optimisation [33, 52, 59] as well as
sparse structures and approximate sparse search [37].
- Statistical signal processing
Although probabilistic methods have played a notable role in extracting high-level information from
signals such as sounds, they have been used less for lower-level signal representation and decomposition.
We have developed new techniques for single- and multi-band amplitude and frequency demodulation
[19, 18, 23, 39, 44], which address the fundamentally ill-posed nature of such problems from a
Bayesian standpoint.
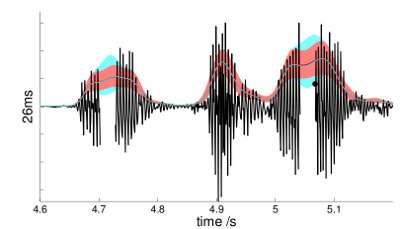
Filling in the envelope of missing segments of a natural signal using PAD.
The red line and region show the estimated envelope and posterior width
using the complete signal; the cyan line and region are estimated from the
signal with drop-outs.
- Dimensionality reduction
Finally, our work on GPFA [28] reflects a more extensive interest in dimensionality reduction in the context of
time-series data—a problem made complicated by uneven sampling and a desire for the simultaneous
characterisation of the embedded manifold and a probabilistical account of the time-series evolution on that
manifold.
Older work
My Ph.D. Thesis on Latent Variable Models for Neural Data Analysis is available from this page. It covers a range
of statistical modelling and data analytic topics, including:
- Relaxation (annealing) schemes for EM and cascading (online) model order selection.
- Identification of spikes from different cells in extracellular recordings.
- Randomly-scaled poisson process models for stimulus-evoked spike trains.
I have also worked on
- MEG denoising and (correlated) source localization (with Srikantan Nagarajan) [52].
- Conditional entropy estimation in low-data categorical experiments.
- Online experimental design [poster].
A selected list of publications appears on the next page.